Двигательный аппарат представлен мускулатурой.
Мышцы, мускулы – органы тела, состоящие из мышечной ткани, способной сокращаться под влиянием нервных импульсов.
Существует три типа мышц: поперечно-полосатые скелетные мышцы, поперечно-полосатая сердечная мышца и гладкие мышцы.
Физиологические свойства мышц:
1) возбудимость — способность возбуждаться при действии раздражителей;
2) проводимость — способность проводить возбуждение;
3) сократимость — способность изменять свою длину или напряжение при возбуждении;
4) растяжимость — способность изменять свою длину под действием растягивающей силы;
5) эластичность — способность восстанавливать свою первоначальную длину после прекращения растяжения.
Сила мышцы определяется максимальным грузом, который мышца может поднять. Мышцы способны совершать работу. Работа мышц определяется произведением величины поднятого груза на высоту подъема. Максимальная работа производится при средних величинах нагрузок. Лабильность мышцы равна 200 — 300 Гц.
При непосредственном раздражении мышцы (прямое раздражение) или опосредованно через иннервирующий ее двигательный нерв (непрямое раздражение) одиночным стимулом возникает одиночное мышечное сокращение, в котором выделяют три фазы: латентный период — время от начала действия раздражителя до начала ответной реакции; фазу сокращения (фаза укорочения) и фазу расслабления.
В естественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, на которые мышца отвечает длительным сокращением. Длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение, называется тетаническим сокращением, или тетанусом. Различают два вида тетануса: зубчатый и гладкий (рис. 6.1).
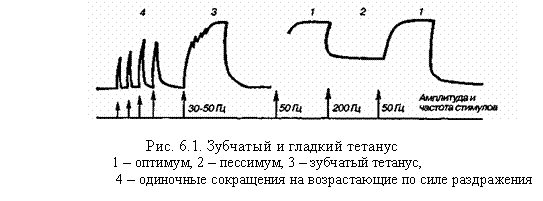
Если каждый последующий стимул поступает к мышце в тот момент, конца она находится в фазе укорочения, то возникает гладкий тетанус, а если в фазу расслабления – зубчатый тетанус.
Амплитуда тетанического сокращения превышает амплитуду одиночного мышечного сокращения. Н.Е. Введенский объяснил это явление фазным изменением возбудимости мышцы, введя понятия об оптимуме и пессимуме частоты раздражения. Оптимум – частота раздражения, при которой каждое последующее раздражение наносится в фазу повышенной возбудимости. Тетанус при этом будет максимальным по амплитуде. Пессимум – частота раздражения, при которой каждое последующее раздражение наносится в фазу пониженной возбудимости. Амплитуда тетануса при этом будет минимальной.
Различают несколько видов мышечных сокращений: изотонический, изометрический и смешанный. При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, если отсутствует сопротивление изменению ее длины. К изотоническому типу сокращений относятся сокращения мышц языка. При изометрическом сокращении длина мышечных волокон остается постоянной, а их напряжение возрастает. Такое сокращение мышцы возникает при попытке поднять чрезмерно большой груз. В естественных условиях сокращения мышц никогда не бывают чисто изотоническими или изометрическими, они имеют смешанный характер, т. е. происходит изменение и длины, и напряжения мышцы.
Механизм мышечного сокращения
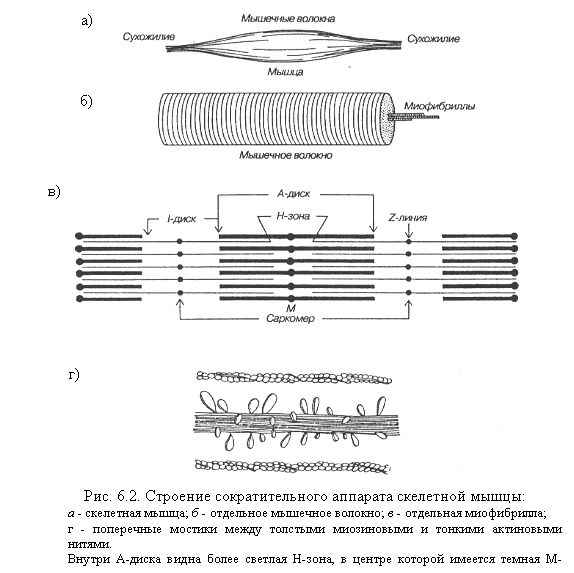
Мышцы состоят из мышечных волокон, а те — из множества тонких нитей — миофибрилл, расположенных продольно (рис. 6.2).
Каждая миофибрилла состоит из нитей сократительных белков актина и миозит. Перегородки, называемые Z-пластинами, разделяют миофибриллы на участки — саркомеры. В саркомере чередуются поперечные светлые и темные полосы. Поперечная исчерченность миофибрилл обусловлена определенным расположением нитей актина и миозина. В центральной части каждого саркомера расположены толстые нити миозина. На обоих концах саркомера находятся тонкие нити актина, прикрепленные к Z-пластинам. Нити миозина выглядят в световом микроскопе как светлые полоски в темном диске, который содержит нити миозина и актина и называется анизотропным, или А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и кажутся светлыми, они называются изотропными, или I-дисками. По их середине проходит темная линия — Z-мембрана. Благодаря такому периодическому чередованию светлых и темных дисков сердечная и скелетная мышцы выглядят поперечно-полосатыми.
В состоянии покоя концы толстых и тонких нитей лишь незначительно перекрываются на уровне А-диска. При сокращении тонкие актиновые нити скользят вдоль толстых миозиновых нитей, двигаясь между ними к середине саркомера. Сами актиновые и миозиновые нити своей длины не изменяют. Миозиновые нити имеют поперечные мостики (выступы) с головками, которые отходят от нити биполярно. Актиновая нить состоит из двух закрученных одна вокруг другой цепочек молекул актина. На нитях актина расположены молекулы тропонина, а в желобках между двумя нитями актина лежат нити тропомиозина. Молекулы тропомиозина в покое располагаются так, что предотвращают прикрепление поперечных мостиков миозина к актиновым нитям.
Во многих местах участки поверхностной мембраны мышечной клетки углубляются в виде трубочек внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-систему). Параллельно миофибриллам и перпендикулярно поперечным трубочкам расположена система продольных трубочек (альфа-система). Пузырьки на концах этих трубочек, в которых сосредоточено основное количество внутриклеточного кальция, подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В состоянии покоя миозиновый мостик заряжен энергией (миозин фосфорилирован), но он не может соединиться с нитью актина, так как между ними находится система из нитей тропомиозина и молекул тропонина. При возбуждении потенциал действия распространяется по мембранам Т-системы внутрь клетки и вызывает высвобождение ионов кальция из альфа-системы. С появлением ионов кальция в присутствии АТФ происходит изменение пространственного положения тропонина — нить тропомиозина сдвигается, и открываются участки актина, присоединяющие миозиновые головки. Соединение головки фосфорилированного миозина с актином приводит к изменению положения мостика (его «сгибанию»), в результате нити актина перемещаются на 1 мм к середине саркомера.
Затем происходит отсоединение мостика от актина. Ритмические прикрепления и отсоединения головок миозина тянут актиновую нить к середине саркомера.
При отсутствии повторного возбуждения ионы кальция закачиваются кальциевым насосом из межфибриллярного пространства в систему саркоплазматического ретикулума. Это приводит к снижению концентрации ионов кальция и отсоединению его от тропонина. Вследствие чего тропомиозин возвращается на прежнее место и снова блокирует активные центры актина. Затем происходит фосфорилирование миозина за счет АТФ, что также способствует временному разобщению нитей. Расслабление мышцы после ее сокращения происходит пассивно — актиновые и миозиновые нити легко скользят в обратном направлении под влиянием сил упругости мышечных волокон, а также сокращения мышц-антагонистов.
Гладкие мышцы
Эти мышцы образуют мышечные слои стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток. Гладкие мышцы разделяются на две основные группы: мультиунитарные и унитарные. Мультиунитарные мышцы функционируют независимо друг от друга, и каждое волокно может иннервироваться отдельным нервным окончанием. Такие волокна обнаружены в ресничной мышце глаза, мигательной перепонке и мышечных слоях некоторых крупных сосудов, к ним относятся мышцы, поднимающие волосы. У унитарных мышц волокна настолько тесно переплетены, что их мембраны могут сливаться, образуя электрические контакты (нексусы). При раздражении одного волокна за счет этих контактов потенциал действия быстро распространяется на соседние волокна. Поэтому, несмотря на то, что двигательные нервные окончания расположены на небольшом числе мышечных волокон, в реакцию вовлекается вся мышца. Такие мышцы имеются в большинстве органов: пищеварительном тракте, матке, в мочеточниках.
Особенностью гладких мышц является их способность осуществлять медленные и длительные тонические сокращения. Медленные, ритмические сокращения гладких мышц желудка, кишечника, мочеточников и других органов обеспечивают перемещение содержимого этих органов. Длительные тонические сокращения гладких мышц обеспечивают функционирование сфинктеров полых органов, которые препятствуют выходу их содержимого.
Гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол, также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосудов влияет на величину их просвета и, следовательно, на уровень кровяного давления и кровоснабжения органов. Важным свойством гладких мышц является их пластичность, т. е. способность сохранять приданную им при растяжении длину. Скелетная мышца в норме почти не обладает пластичностью. При удалении растягивающего груза скелетная мышца быстро укорачивается, а гладкая с становится растянутой. Высокая пластичность гладких мышц имеет большое значение для нормального функционирования полых органов. Например, пластичность мышц мочевого пузыря по мере его наполнения предотвращает избыточное повышение давления.
Сильное и резкое растяжение гладких мышц вызывает их сокращение, что обусловлено нарастающей при растяжении деполяризацией клеток, которая обеспечивает автоматику гладкой мышцы. Такое сокращение играет важную роль в авторегуляции тонуса кровеносных сосудов, а также способствует непроизвольному опорожнению переполненного мочевого пузыря в тех случаях, когда нервная регуляция отсутствует в результате повреждения спинного мозга.
В гладких мышцах тетаническое сокращение возникает при низкой частоте стимуляции. Особенностью гладких мышц является их высокая чувствительность к медиаторам. При нанесении ацетилхолина на препарат мышцы толстой кишки частота потенциала действия возрастает. Вызываемые ими сокращения сливаются, образуется почти гладкий тетанус. Чем выше частота потенциала действия, тем сильнее сокращение. Норадреналин, напротив, гиперполяризует мембрану, снижая частоту потенциала действия и величину тетануса.
Возбуждение гладкомышечных клеток вызывает повышение концентрации кальция в саркоплазме, что активирует сократительные структуры. Так же как сердечная и скелетная мышцы, гладкая мышца расслабляется при снижении концентрации ионов кальция. Расслабление гладких мышц происходит медленнее, так как удаление ионов кальция замедлено.